
Leatherbacks, Light, and the Comparative Study of Seafinding Behavior
Samantha Trail and Michael Salmon Florida Atlantic University .

Sea turtles are unique in that they are large, long-lived species, but they produce a large number of offspring with very few surviving to reach maturity themselves. Assuming ideal nest conditions, that treacherous struggle to survive begins almost immediately. As hatchlings, sea turtles emerge from the sand with their clutch mates, experiencing the beach environment outside of the buried egg chamber for the very first time. The immediate objective: get to the ocean as quickly and accurately as possible.
This journey from the nest to the ocean is called “seafinding” – an innate behavior under intense selection pressure requiring a consistent, reliable cue. It is now well documented that hatchlings of all species accomplish this feat by crawling in the direction of the lowest and brightest horizon. Regardless of lunar phase or position under natural lighting conditions, that lowest, brightest horizon will be toward the sea (Figure 1). The turtles then use a “positive phototaxis” (attraction to the brightest location) to crawl in that direction.
However when animals respond to visual cues, they favor some wavelengths over others - those that are particularly important for promoting adaptive behavior. For example, foraging honeybees use the pattern of ultra-violet (UV)
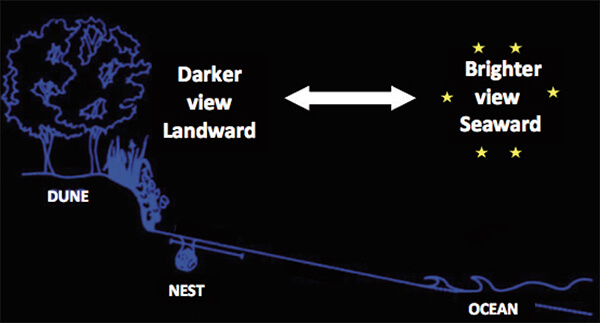
Figure 1. A view of the nesting beach at night. Hatchlings emerging from the nest will use vision to scan their environment before crawling toward the sea. The view toward land (here, a dune) will be darker as light is absorbed by vegetation. In contrast, light from the stars (and moon, if present) is reflected from the water surface making the view toward the sea brighter. This difference tells the hatchlings where the ocean is located, even if they can’t directly see it from the nest site, and triggers a positive phototaxis.
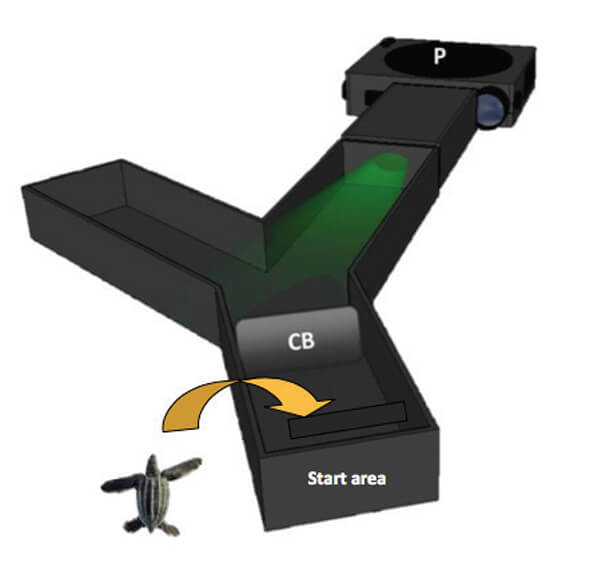
Figure 2. A Y-maze is used to reproduce a light intensity difference, similar to the one found at the nesting beach. A projector (P) is used to present a light stimulus in one of the Y-maze arms; the other arm remains dark. If hatchlings can detect the light, they will crawl into the illuminated arm. When the light intensity is reduced to a level that they can’t see, hatchlings are equally likely to crawl into either arm. That difference enables investigators to obtain wavelength phototaxis thresholds. CB is the clear barrier that keeps the turtle in the start area until the light is presented.
That exquisite sensitivity meant they could easily distinguish between differences in light intensity in a landward vs. seaward direction, even on the darkest of evenings.
Those results piqued our interest into whether leatherback hatchlings, which are distantly related to both green turtles and loggerheads, had evolved a similar or different pattern of sensitivity. Leatherbacks diverged from their hard-shelled relatives ~ 100 million years ago and are also very unique turtles! They are exclusively oceanic, deep-diving residents throughout their life, as opposed to mature loggerheads and green turtles that live primarily in shallow coastal waters. So, though visual cues are required for seafinding by all marine turtle hatchlings, those ecological differences and the long evolutionary separation between the two groups provide the potential for significant differences in how they accomplish their crawl to the ocean. Our objective was to find out the extent to which that potential was realized.
We used the same methods and obtained turtles from the same nesting beaches as Celano’s group to measure leatherback spectral sensitivities (Figure 3).
To investigate this question, we headed to the beach to observe and compare the natural paths of loggerhead and leatherback hatchlings crawling under full (or nearly full) and new moon conditions. Though loggerheads showed similar seafinding paths under full or new moon conditions, the same could not be said of the leatherbacks. Two major differences were revealed: under new moon conditions, leatherbacks (1) showed considerably more circling during their crawl (Figure 4) and (2) their crawling orientation angles under those conditions were significantly more variable (Figure 5). We think that outcome is related to their light sensitivity. Under new moon conditions, leatherback phototaxis thresholds are only slightly below background light levels (red lines in Figure 3), so distinguishing between landward and seaward light intensities might be more challenging. It’s likely, then, that circling follows in an attempt to make that intensity distinction clearer. Even so, leatherbacks usually manage to find the sea during new moon. It’s just more difficult!
Our findings should be of interest to conservation managers as they also suggest that leatherback seafinding, whether by hatchlings or adult females after nesting, will be more prone to disruption by artificial light sources, including sky glow that currently poses a serious threat worldwide to all marine turtles at nesting beaches. We suspect that as a consequence, leatherback hatchlings at urban beaches may fail to locate the sea more often than the hard-shelled hatchlings. Given that various leatherback populations worldwide are in serious decline, further research into this problem is certainly appropriate.
wavelengths shown by flowers to identify those that are currently producing nectar. Pit vipers, hunting at night, are sensitive to the longer, infrared wavelengths that generate heat as heat also radiates from their warm-blooded mammalian prey; that “heat image” directs the snake’s venomous strike. Similarly, we expect sea turtle hatchlings to be particularly sensitive to the light wavelengths present at the beach at night, especially if they can be used to distinguish between a landward and an oceanic view. But, how can that sensitivity best be measured? The solution is to create an environment in which, once again, hatchlings need to choose between two spatially separate locations, one visually darker than the other, so that a positive phototaxis response is released.
Master’s graduate Lisa Celano and her colleagues were the first to use this technique to measure light spectral sensitivities in loggerhead and green turtle hatchlings, two species of “hard-shelled” marine turtle. They used a Y-maze with light wavelengths projected from one arm but not the other (Figure 2), and sequentially presented individual light wavelengths between 340 and 600 nm (near-UV to orange). They knew from measurements made at the beach that these light energies were present at night. Initially, the light stimulus was bright enough to induce all the turtles to crawl into the illuminated arm. That intensity was then systematically reduced until the preference for the illuminated arm was eliminated. The lowest light intensity that still induced a group of hatchlings to crawl into the illuminated arm was recorded as the “phototaxis threshold”, or sensitivity measure, for that particular wavelength.
The data from that study (Figure 3) revealed three trends. First, loggerheads and green turtles had almost identical sensitivities, a sure sign of what biologists call “convergent evolution.” That finding meant that the two species, even though separately evolved for ~ 63 million years, had settled on the same solution to the seafinding problem. Second, both species were most sensitive to the shorter (<500 nm) wavelengths. Third, both species were up to 100 times more sensitive to those wavelengths than to the background illumination presented by the stars or moonlight at the beach (Figure 1).
Our results provide support for the idea that seafinding was accomplished in leatherbacks through somewhat different, or “divergent,” means. Like loggerheads and green turtles, leatherbacks were most sensitive to shorter wavelengths (<500 nm) but leatherbacks were consistently less sensitive to all the tested wavelengths than their hard-shelled relatives. This difference in overall sensitivity was profound, as it meant that leatherbacks required upwards of 10 times more light at nearly all the utilized wavelengths to direct their crawl to the sea.
Intrigued by these differences, we explored another question: How might this reduction in sensitivity translate into behavioral differences during seafinding on the beach? Previous observations by other naturalists (Archie Carr and Nicholas Mrosovsky) suggest that one peculiar seafinding behavior occurred more often in leatherback hatchlings than others: circling. More specifically, orientation circles are characteristic when a hatchling (or nesting females) during their crawl does not head in a straight direction but instead makes brief, tight circles, and less often as a larger, sweeping path. Was that circling somehow related to differences in light sensitivity between the two groups of marine turtles?
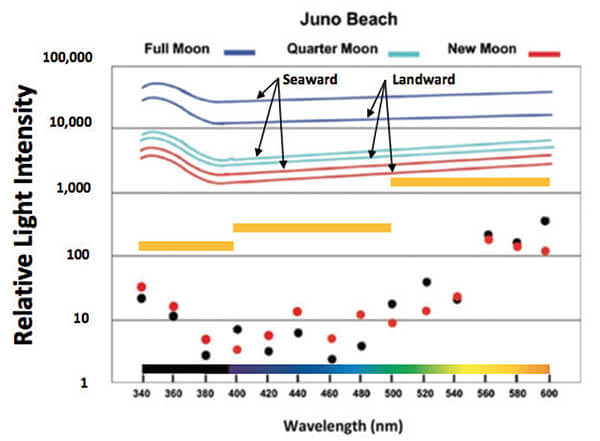
Figure 3. Phototaxis thresholds for Green Turtles () and Loggerheads () plotted relative to the mean thresholds for Leatherbacks () between 340 - 400 nm, 400 - 500 nm, and 500 - 600 nm. Thresholds are shown relative to background lighting measured during a full moon, quarter moon and new moon (paired colored lines, above). All three species can detect the presence of light well below background, including the ultraviolet range (below 400 nm) that humans do not see but that is present at night. The two hard-shelled species are between 10 - 100 x more sensitive to light than leatherbacks and under all conditions should easily distinguish between the darker view toward land and the brighter view toward the sea. However, leatherbacks are much less sensitive and especially during new moon, when thresholds are not much lower than background, might find that task more difficult.

Figure 4. A comparison of leatherback hatchling crawling tracks during seafinding under full moon (blue) and new moon (red) lighting conditions. Hatchlings were released from the nest site and allowed to crawl unencumbered to the wrack line on the beach. During new moon, leatherback hatchling orientation was less accurate and contained several bouts of circling, as seen in this track record.
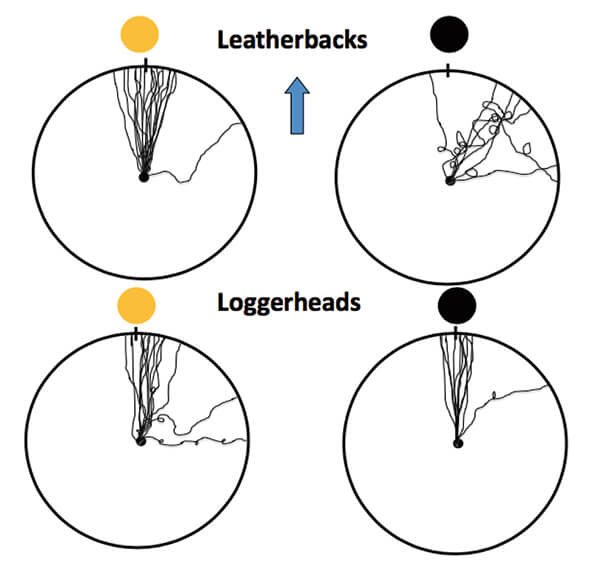
Figure 5. Diagrams showing how leatherback (top row) and loggerhead (bottom row)
hatchlings oriented during their crawl between the nest and the location of the ocean,
as indicated by the blue arrow, on nights when exposed to a full moon (left column)
or a new moon (right column). Each line represents the track of one turtle. Leatherback
accuracy declines during new moon but not during full moon when light intensity cues
are easier to distinguish from background (see Figure 3). A similar decline does not occur
in loggerheads, probably because of their greater sensitivity to nocturnal lighting.
hatchlings oriented during their crawl between the nest and the location of the ocean,
as indicated by the blue arrow, on nights when exposed to a full moon (left column)
or a new moon (right column). Each line represents the track of one turtle. Leatherback
accuracy declines during new moon but not during full moon when light intensity cues
are easier to distinguish from background (see Figure 3). A similar decline does not occur
in loggerheads, probably because of their greater sensitivity to nocturnal lighting.

Helping Sea Turtles Survive for 39 Years
A NON-PROFIT ORGANIZATION
State of Florida Registration Number CH-2841 | Internal Revenue Code 501 (c) (3)
Web Design & Development by Web Expressions, LLC